22/04/2020
Los capilares sistémicos son la red más fina y extensa de vasos sanguíneos en nuestro cuerpo, actuando como el principal escenario para el intercambio de sustancias entre la sangre y los tejidos. Aunque a menudo invisibles a simple vista, su función es absolutamente crucial para la vida, permitiendo que las células reciban oxígeno y nutrientes vitales, y que se eliminen los productos de desecho. Comprender la ultraestructura de sus paredes y cómo regulan la permeabilidad es fundamental para desentrañar los misterios de la fisiología humana.
- La Arquitectura Microscópica de los Capilares: Más Allá de lo Visible
- Componentes Celulares Clave y su Función en la Permeabilidad
- El Glicocálix Endotelial: El Filtro Maestro
- Mecanismos de Transporte a Través de la Pared Capilar
- Permeabilidad Diferencial para Diversas Sustancias
- La Base Ultraestructural de la Permeabilidad: Fenestraciones y Hendiduras Intercelulares
- Preguntas Frecuentes (FAQs)
- ¿Cuál es la función principal de los capilares sistémicos?
- ¿Qué diferencia a un capilar continuo de un capilar fenestrado?
- ¿Qué es el glicocálix y por qué es importante?
- ¿Cómo se transportan el oxígeno y el dióxido de carbono a través de la pared capilar?
- ¿Qué son las uniones estrechas en los capilares y cuál es su papel?
La Arquitectura Microscópica de los Capilares: Más Allá de lo Visible
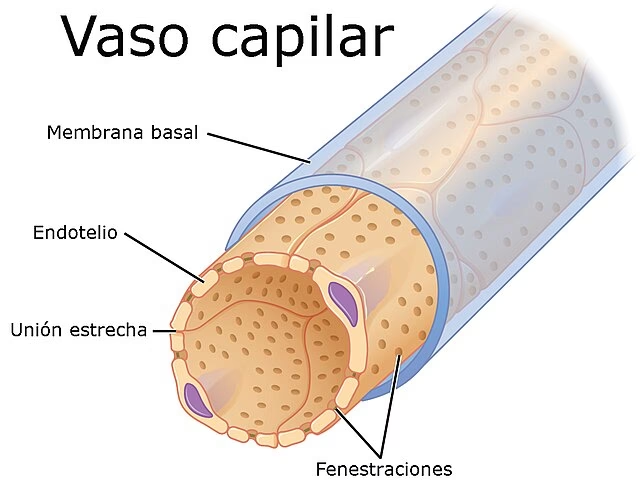
A primera vista, las paredes capilares parecen increíblemente simples, compuestas por una sola capa de células aplanadas conocidas como células endoteliales. Sin embargo, esta simplicidad esconde una complejidad funcional asombrosa. Observaciones con microscopía electrónica han revelado que el endotelio capilar no es uniforme, sino que se presenta en dos tipos principales, cada uno adaptado a las necesidades específicas de los tejidos que irriga.
Endotelio Continuo: La Barrera Rigurosa
El endotelio continuo es el tipo más común y se encuentra en microvasos de la piel, los músculos, los pulmones y los tejidos conectivos. En este tipo de capilares, las células endoteliales están estrechamente unidas entre sí por 'uniones estrechas', formando una capa ininterrumpida que rodea una membrana basal continua. Estas uniones actúan como una barrera selectiva, controlando rigurosamente qué sustancias pueden pasar de la sangre a los tejidos y viceversa. La integridad de las membranas plasmáticas de estas células se mantiene incluso en áreas donde se aplanan, reduciendo su espesor a menos de 0.1 micrómetros.
Endotelio Fenestrado: La Puerta al Intercambio Rápido
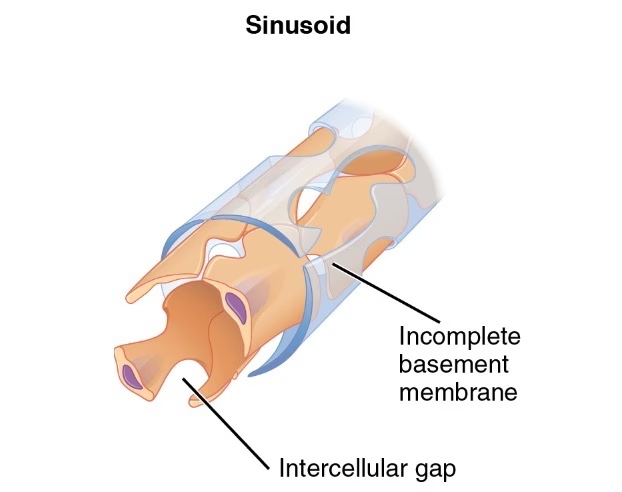
En contraste, el endotelio fenestrado se encuentra en microvasos asociados con epitelios secretores y absorbentes, como los capilares de las vellosidades de la mucosa intestinal y los capilares glomerulares y peritubulares del riñón. Las paredes de estos microvasos también están formadas por una única capa continua de células endoteliales unidas por uniones estrechas y rodeadas por una membrana basal continua. Sin embargo, la característica distintiva son las 'fenestraciones' o 'ventanas', aberturas circulares de 40 a 70 nanómetros de diámetro que parecen penetrar las áreas atenuadas de las células. En la mayoría de los casos, estas fenestraciones están cerradas por un delgado diafragma denso en electrones, que se organiza como una serie de radios anchos con un 'centro' central. Estas fenestraciones permiten un intercambio mucho más rápido y eficiente de fluidos y solutos, vital para la función de órganos como los riñones y el intestino.
Tabla Comparativa: Endotelio Continuo vs. Fenestrado
| Característica | Endotelio Continuo | Endotelio Fenestrado |
|---|---|---|
| Localización Principal | Piel, músculo, pulmón, tejido conectivo | Intestino, riñón (glomérulos, peritubulares), glándulas endocrinas |
| Estructura de la Pared | Capa continua de células endoteliales | Capa continua de células endoteliales con poros (fenestraciones) |
| Uniones Intercelulares | Uniones estrechas (tight junctions) muy compactas | Uniones estrechas, pero con fenestraciones |
| Permeabilidad | Baja a moderada, altamente selectiva | Alta, permite paso rápido de fluidos y solutos pequeños |
| Función Principal | Intercambio controlado de gases, nutrientes y desechos | Secreción, absorción y filtración rápida |
Componentes Celulares Clave y su Función en la Permeabilidad
Más allá de la distinción entre endotelio continuo y fenestrado, las células endoteliales poseen una serie de componentes internos que orquestan su función de barrera y transporte.
Caveolas y Tráfico Vesicular
Una característica ultraestructural dominante en las células endoteliales es la gran cantidad de pequeñas vesículas endoplasmáticas o caveolas. Antes se pensaba que cada vesícula era una entidad separada, pero ahora se sabe que la mayoría se organizan en grupos fusionados que se comunican con las superficies luminal o abluminal de las células. Estas vesículas, que miden aproximadamente 70 nm de diámetro y están recubiertas de proteínas como la caveolina, desempeñan un papel en el transporte de sustancias a través de la célula (transcitosis), aunque las cadenas de vesículas fusionadas que forman canales transcelulares son relativamente raras en el endotelio no estimulado.
El Citoesqueleto: Soporte y Dinamismo
El citoesqueleto endotelial, aunque menos conspicuo que las vesículas, es una red compleja de tres tipos de fibras: actina polimerizada, microtúbulos e filamentos intermedios. Las fibras de actina se presentan en una red cortical bajo la superficie celular, como fibras de estrés alineadas con las fuerzas de cizallamiento, y como filamentos asociados a las uniones (JAF). Estas estructuras no solo proporcionan soporte mecánico, sino que también son dinámicas, respondiendo a estímulos y participando en cambios de la forma celular y la transmisión de fuerzas mecánicas, lo que puede influir en la permeabilidad de la pared capilar.
Uniones Intercelulares y Hendiduras: Las Rutas Paracelulares
Las células endoteliales se unen entre sí en regiones de unión donde existe una hendidura de 15 a 20 nm de ancho. Sin embargo, en ciertos puntos, esta hendidura se estrecha, y las membranas externas de las células adyacentes parecen fusionarse, formando las 'uniones estrechas'. Estas uniones, compuestas por hebras de moléculas, son cruciales para la permeabilidad. Aunque impiden la difusión de iones grandes, presentan 'roturas' ocasionales que actúan como vías para pequeños solutos hidrofílicos a través de la barrera endotelial. La complejidad y la frecuencia de estas roturas en las uniones estrechas varían entre los diferentes lechos capilares y son un determinante clave de su permeabilidad.
El Glicocálix Endotelial: El Filtro Maestro
Una característica ultraestructural de vital importancia para la permeabilidad microvascular es el glicocálix del endotelio. Esta capa de proteínas glicosiladas cubre la superficie luminal de las células endoteliales, incluyendo las fenestraciones, y se extiende desde 30-50 nm hasta 1-3 micrómetros hacia el lumen del vaso. Identificado inicialmente mediante microscopía electrónica con tinciones catiónicas, el glicocálix ha sido reconocido como un componente fundamental de la barrera capilar.
Composición y Estructura
El glicocálix está compuesto por una variedad de moléculas, incluyendo proteoglicanos y glicoproteínas. La hipótesis de que actúa como un filtro molecular ha sido ampliamente respaldada. Análisis de imágenes recientes sugieren una estructura cuasiperiódica, un enrejado regular con espacios internos de aproximadamente 8 nm de lado, lo que es consistente con su papel como filtro molecular. Esta intrincada red filamentosa es la que, en última instancia, determina la exclusión de macromoléculas del paso a través de la pared capilar.
Función de Barrera: El Ultrafiltro
El glicocálix actúa como el principal ultrafiltro para las macromoléculas en la mayoría de las paredes microvasculares. Su presencia restringe el movimiento de moléculas grandes, como las proteínas plasmáticas, impidiendo que se escapen al espacio intersticial. De hecho, experimentos han demostrado que la alteración del glicocálix, por ejemplo, con enzimas, puede duplicar la permeabilidad hidráulica de los capilares, lo que subraya su papel crítico en el mantenimiento de la homeostasis de fluidos y proteínas entre la sangre y los tejidos.
Mecanismos de Transporte a Través de la Pared Capilar
El transporte de fluidos y solutos a través de las paredes capilares es un proceso pasivo impulsado por diferencias en la energía potencial de las soluciones a cada lado de la membrana. Los dos mecanismos principales son la convección y la difusión.
Transporte Pasivo: Convección y Difusión
- Convección: Se refiere al flujo masivo de soluciones y los solutos dentro de ellas, impulsado por diferencias de presión hidrostática. La facilidad con la que el fluido fluye a través de la membrana se describe por la conductividad hidráulica o permeabilidad hidráulica (Lp). Si el flujo de soluto es más restringido que el del agua, una fracción de las moléculas de soluto es 'rechazada' o 'reflejada' por la membrana, lo que se describe por el coeficiente de reflexión (σf).
- Difusión: Es el mecanismo más importante para el transporte de moléculas pequeñas a través de las paredes microvasculares. Resulta del movimiento aleatorio de las moléculas debido a su energía térmica. La difusión neta ocurre desde regiones de alta concentración a regiones de baja concentración, y se describe por la ley de Fick. La permeabilidad difusional (Pd) de la membrana al soluto se define en términos del flujo neto del soluto, la diferencia de concentración a través de la membrana y el área de la membrana.
Coeficientes de Permeabilidad: Lp, Pd y σ
Estos coeficientes cuantifican la facilidad con la que las sustancias atraviesan la pared capilar. El coeficiente de permeabilidad hidráulica (Lp) describe el flujo de fluido, el coeficiente de permeabilidad difusional (Pd) describe el flujo de solutos por difusión, y el coeficiente de reflexión (σ) describe el grado de rechazo de un soluto por la membrana, tanto por convección (σf) como por presión osmótica (σd). Para un soluto ideal, σf es igual a σd. Los valores de σ varían de 1.0 (rechazo total) a 0 (ningún rechazo), e incluso pueden ser negativos en casos muy específicos.
Permeabilidad Diferencial para Diversas Sustancias
La capacidad de las paredes capilares para permitir el paso de diferentes tipos de moléculas varía drásticamente según sus propiedades.
Solutos Lipofílicos: Paso Directo y Rápido
Los solutos que pueden disolverse significativamente en lípidos, como el oxígeno (O2), el dióxido de carbono (CO2) y los anestésicos generales, tienen muy altas permeabilidades microvasculares. Se asume que estas moléculas difunden directamente a través de toda la pared microvascular, incluyendo las membranas celulares endoteliales. Su transporte entre la sangre y los tejidos es tan rápido que, bajo condiciones fisiológicas, su velocidad de intercambio está limitada por el flujo sanguíneo, no por la barrera capilar misma. Esto significa que la pared capilar no representa una barrera significativa para estas sustancias.
Solutos Hidrofílicos Pequeños: Rutas Restringidas
La permeabilidad a las moléculas hidrofílicas (que se disuelven en agua) depende en gran medida de su tamaño molecular. Para moléculas pequeñas (como iones de Na+, K+, Cl- o glucosa, aminoácidos, urea), los coeficientes de permeabilidad (Pd) disminuyen bruscamente a medida que aumenta el tamaño molecular hasta un cierto punto (aproximadamente el tamaño de la albúmina sérica). Esta disminución más rápida de Pd de lo que se esperaría solo por la disminución del coeficiente de difusión en el agua, sugiere una 'difusión restringida' a través de canales llenos de agua cuyas dimensiones son comparables a los diámetros de las moléculas que difunden. Las variaciones en Lp y Pd para estos solutos entre diferentes lechos microvasculares (por ejemplo, los capilares mesentéricos son 20-30 veces más permeables a Na+ y K+ que los del músculo esquelético) se correlacionan fuertemente, lo que sugiere que las mismas vías extracelulares (hendiduras intercelulares) sirven tanto para el intercambio rápido de solutos hidrofílicos pequeños como para la mayoría de los movimientos netos de fluido. Aunque las acuaporinas (canales de agua, como AQP-1) contribuyen al flujo de agua, su aporte a la Lp total de la mayoría de los vasos de intercambio es relativamente pequeño (5-10%).
Macromoléculas: La Barrera del Glicocálix
Las macromoléculas, como las proteínas plasmáticas (por ejemplo, la albúmina sérica), tienen valores de coeficientes de reflexión (σ) muy altos (0.8-0.999), lo que indica que son fuertemente rechazadas por la barrera capilar. A pesar de las grandes variaciones en la Lp entre diferentes lechos microvasculares, los coeficientes de reflexión para las macromoléculas son notablemente similares en vasos no dañados. Esto sugiere que los canales o poros responsables de la Lp en diferentes tejidos restringen el paso de la albúmina en una medida similar. La principal estructura responsable de este filtrado molecular para las macromoléculas es el glicocálix luminal del endotelio, que actúa como el ultrafiltro esencial.
La Base Ultraestructural de la Permeabilidad: Fenestraciones y Hendiduras Intercelulares
La comprensión detallada de la ultraestructura capilar nos permite identificar las vías exactas por las que el agua y los solutos se mueven.
Capilares Fenestrados: Eficiencia en el Intercambio
En los capilares con endotelio fenestrado, las fenestraciones son las principales responsables de los altos valores de Lp y Pd para pequeños solutos hidrofílicos. Aunque las fenestraciones mismas son demasiado grandes para explicar el cribado de macromoléculas, el glicocálix, junto con los diafragmas fenestrales y la membrana basal, es el responsable de este filtrado. Las variaciones en la Lp de los vasos fenestrados se correlacionan con el número de fenestraciones por unidad de área del endotelio.
Capilares Continuos: La Ruta Tortuosa de las Hendiduras
En los vasos con endotelio continuo, las vías principales para el agua y los solutos hidrosolubles pequeños se encuentran en las hendiduras intercelulares. Investigaciones detalladas han demostrado que, si bien las uniones estrechas son la principal barrera, existen 'roturas' ocasionales en las hebras de estas uniones que permiten un paso tortuoso a través de la hendidura desde el lumen del vaso hasta la membrana basal. La frecuencia y las dimensiones de estas roturas son suficientes para explicar la permeabilidad hidráulica observada. La complejidad de la arquitectura de las uniones estrechas dentro de las hendiduras intercelulares determina las amplias variaciones de permeabilidad entre diferentes tipos de capilares continuos (por ejemplo, los capilares mesentéricos son más permeables que los cardíacos, y estos más que los del músculo esquelético, debido a una menor complejidad de las uniones y, por lo tanto, vías más directas).
En resumen, los capilares sistémicos son maravillas de la ingeniería biológica. Su capacidad para regular el paso de sustancias, desde gases lipofílicos que atraviesan las membranas celulares directamente, hasta pequeñas moléculas hidrofílicas que se abren camino a través de uniones intercelulares especializadas, y macromoléculas que son cuidadosamente filtradas por el glicocálix, es un testimonio de la precisión con la que nuestro cuerpo mantiene la homeostasis.
Preguntas Frecuentes (FAQs)
¿Cuál es la función principal de los capilares sistémicos?
La función principal de los capilares sistémicos es facilitar el intercambio de oxígeno, nutrientes, hormonas y otros compuestos esenciales desde la sangre hacia los tejidos, y la eliminación de dióxido de carbono y productos de desecho desde los tejidos hacia la sangre. Actúan como el puente entre las arteriolas y las vénulas.
¿Qué diferencia a un capilar continuo de un capilar fenestrado?
La principal diferencia radica en la estructura de sus células endoteliales. Los capilares continuos tienen células endoteliales estrechamente unidas sin poros, formando una barrera más restrictiva. Los capilares fenestrados, en cambio, poseen 'ventanas' o poros (fenestraciones) en sus células endoteliales que permiten un intercambio más rápido de fluidos y solutos, siendo comunes en órganos con alta necesidad de filtración o secreción.
¿Qué es el glicocálix y por qué es importante?
El glicocálix es una capa de proteínas glicosiladas que recubre la superficie luminal (interna) de las células endoteliales capilares. Es crucial porque actúa como el principal 'filtro molecular' para las macromoléculas, como las proteínas plasmáticas. Impide que estas moléculas grandes se escapen de la sangre hacia los tejidos, manteniendo así el equilibrio osmótico y de fluidos.
¿Cómo se transportan el oxígeno y el dióxido de carbono a través de la pared capilar?
El oxígeno y el dióxido de carbono son solutos lipofílicos, lo que significa que pueden disolverse fácilmente en las membranas lipídicas. Por lo tanto, difunden directamente a través de toda la pared de las células endoteliales capilares. Su transporte es tan eficiente que su velocidad de intercambio está limitada por el flujo sanguíneo, no por la barrera capilar en sí.
¿Qué son las uniones estrechas en los capilares y cuál es su papel?
Las uniones estrechas son puntos donde las membranas de las células endoteliales adyacentes se fusionan, formando una barrera continua. En los capilares continuos, estas uniones contienen 'roturas' ocasionales que actúan como las principales vías para el paso de agua y solutos hidrofílicos pequeños. La complejidad y la frecuencia de estas roturas determinan la permeabilidad de un capilar continuo a estas sustancias.
Si quieres conocer otros artículos parecidos a ¿Qué son los Capilares Sistémicos y Cómo Funcionan? puedes visitar la categoría Cabello.